- MÉIOSE
- MÉIOSELa reproduction d’un être vivant peut s’effectuer selon deux voies. L’une, dite végétative , constitue la voie de reproduction conforme de l’organisme ou d’une partie de celui-ci, qui peut être représentée par une seule cellule. L’autre voie, dite sexuée , permet la réunion (par fusion de cellules spécialisées dites sexuelles ou gamètes) de deux patrimoines génétiques issus de deux clones cellulaires ou d’individus distincts, les parents. La reproduction sexuée permet donc un brassage constant des potentialités héréditaires à l’intérieur d’une même espèce.Chaque réunion entre deux gamètes de sexe opposé, ou fécondation, produit une cellule nommée zygote, ou, plus couramment, œuf. Elle est diploïde, c’est-à-dire qu’elle contient chacun des chromosomes caractéristiques de l’espèce en deux exemplaires (2 n ) puisqu’ils proviennent des deux parents.Il est donc indispensable que, préalablement à la formation des gamètes de l’individu diploïde, se produise toujours un mécanisme régulateur qui restitue aux cellules sexuelles leur contenu chromosomique haploïde.Le mécanisme de ce processus, que l’on appelle la méiose , est un enchaînement de deux divisions cellulaires accompagnées d’une seule duplication du matériel chromosomique. La cellule diploïde (2 n ) qui subit la méiose donne naissance à quatre cellules haploïdes (n ). Non seulement ce mécanisme connu chez tous les Eucaryotes préserve le nombre de chromosomes de l’espèce, mais il permet l’apparition de nouveaux types génétiques.Quant au déterminisme de la méiose, il reste en grande partie inconnu; son induction comme son déroulement semblent conditionnés par des facteurs génétiques, hormonaux, ainsi que par le milieu [cf. REPRODUCTION].Après avoir dégagé les aspects cytologiques essentiels de l’histoire des chromosomes au cours des deux divisions méiotiques, on en abordera les conséquences génétiques.Aspects cytologiquesL’histoire des chromosomes au cours de la méiose est remarquablement constante chez tous les Eucaryotes (alors que rien de tel n’intervient dans le cas des Procaryotes). La méiose est composée de deux divisions successives, mais les chromosomes – donc l’ADN, c’est-à-dire l’acide désoxyribonucléique dont ils sont formés – ne subissent qu’une seule duplication, effectuée au moins en grande partie lorsque commence la prophase de première division. Dans certains cas (champignons Ascomycètes), on a montré qu’elle avait lieu avant même la fusion des deux noyaux haploïdes.Pendant la prophase de la première division, les chromosomes s’associent par paires, chaque chromosome étant lui-même formé (duplication) de deux chromatides. À ce stade se produisent les échanges entre le chromosome venant de la mère et celui qui provient du père. Les chromosomes de chaque paire migrent ensuite à un pôle opposé, et les deux noyaux formés à la fin de la première division contiennent ainsi un jeu haploïde (n ) de chromosomes mais dont chacun est clivé en deux chromatides. Comme il n’y a pas de nouvelle duplication chromosomique avant la deuxième division, les deux chromatides de chaque chromosome se séparent, et on aboutit ainsi à la formation de quatre noyaux possédant chacun un jeu haploïde de chromosomes. Ce comportement chromosomique est semblable dans les deux sexes mais, dans la lignée femelle des animaux et celle des végétaux supérieurs, un seul des quatre noyaux haploïdes subsiste. Après la première division, un des deux noyaux est expulsé dans une petite cellule, ou globule polaire (chez les animaux), et ne se divisera plus. Il en sera de même pour un des noyaux de deuxième division: trois des quatre produits de la méiose sont ainsi éliminés.Prophase de première divisionCette phase, longue et totalement différente d’une prophase mitotique, a été divisée en cinq stades. Le leptotène (du grec leptos : mince) correspond à l’individualisation des chromosomes (fig. 1 a) sous forme de longs filaments jalonnés d’épaississements très colorables appelés chromomères. Chaque chromosome est déjà dupliqué et donc formé de deux chromatides. Les extrémités chromosomiques ou télomères sont attachées à l’enveloppe nucléaire, le plus souvent (surtout chez les animaux) dans une zone particulière face au centrosome. Cet arrangement dit «en bouquet» est conservé durant les deux premiers stades.Le zygotène (zygos : paire) est caractérisé par l’appariement progressif (fig. 1 b, flèches) des chromosomes homologues (l’un venant du père, l’autre de la mère). On connaît mal la nature des forces physico-chimiques qui permettent cet appariement spécifique (cf. infra ), mais ce processus, souvent appelé synapsis , est l’un des phénomènes essentiels de la méiose. Chaque paire de chromosomes homologues ainsi formée constitue un bivalent : on observe donc n bivalents par noyau. Les chromosomes sexuels (X et Y chez l’homme), seulement partiellement homologues, ne s’apparient que dans la région homologue, et, s’ils sont entièrement hétérologues, ils ne s’apparient pas.Au pachytène (pakhus : épais), les chromosomes appariés sur toute leur longueur s’épaississent et se raccourcissent (fig. 2). Les télomères se détachent de l’enveloppe nucléaire, et l’arrangement en bouquet disparaît.Lorsque les deux homologues de chaque paire commencent à se séparer, le stade est appelé diplotène (diploos : double). Les chromosomes sont clivés depuis le leptotène en deux chromatides, sauf au niveau du centromère, mais ce n’est qu’au diplotène que ce dédoublement est facilement visible: le bivalent est alors souvent décrit comme une tétrade . La séparation n’est pas totale car les paires de chromatides restent associées en un ou plusieurs points (fig. 3). Ces régions, auniveau desquelles une des chromatides de chaque paire semble changer de partenaire (fig. 3 b) pour aller s’apparier avec l’autre chromatide homologue, sont appelées chiasma . Leur nombre est variable d’un chromosome à l’autre (le nombre moyen est fonction de la longueur du chromosome, fig. 3) et, pour un même chromosome, d’une méiose à l’autre. Le chiasma n’est pas un simple enroulement des deux chromosomes mais bien le résultat d’un échange. En effet, lorsqu’on peut différencier les deux homologues (par exemple, perte d’une extrémité pour l’un), on observe toujours que les portions de chromatides appariées après le chiasma sont de taille identique , figure qui ne peut s’expliquer que par l’occurrence d’un échange entre deux des quatre chromatides homologues. Cette interprétation est confirmée par des expériences de marquage radioactif de l’ADN. Lorsqu’un précurseur radioactif est fourni pendant la synthèse d’ADN préméiotique, chaque chromosome dupliqué possède une chromatide radioactive et une chromatide non radioactive. À la fin du diplotène, les chromatides présentent une alternance de segments marqués (radioactifs) et de segments non marqués avec une distribution complémentaire. Cet échange précis et parfaitement réciproque est appelé crossing-over (cf. infra ). Bien qu’il paraisse clair que le chiasma soit une conséquence cytologique du crossing-over, son mécanisme est encore mal compris. Dans les ovocytes, le diplotène peut être de longue durée: chez la femme, il commence au troisième mois de la vie fœtale et s’arrête au plus tôt à la puberté.À la diacinèse (dia : à travers; kinesis: mouvement ), la séparation des homologues s’accentue en même temps que les chromosomes se raccourcissent fortement (fig. 6).Complexe synaptonémal, mécanismes d’appariement et recombinaison génétiqueLe microscope électronique a révélé la formation d’une structure protéique spécifique de l’appariement: le complexe synaptonémal (fig. 4). Au pachytène, chaque chromosome est associé avec un élément dense appelé élément latéral dont la longueur représente environ 1 p. 100 de celle de son ADN. Lorsque l’appariement est réalisé, un élément central apparaît, qui relie les éléments latéraux. La formation de cette structure tripartite est, chez les Eucaryotes, un phénomène aussi universel que la méiose elle-même.La reconstitution des chromosomes à partir de noyaux coupés en série a permis d’analyser l’évolution du complexe durant la prophase chez plusieurs espèces (fig. 5). L’élément latéral est mis en place dès le leptotène mais n’apparaît continu qu’au zygotène. Ses deux extrémités sont ancrées à la membrane interne de l’enveloppe nucléaire. L’appariement est consolidé par la mise en place de l’élément central. Ce dernier est formé à égale distance des deux éléments latéraux lorsque les homologues sont à une distance de 100 à 150 nm. Cette distance est rigoureusement égale dans tous les bivalents et a été trouvée constante dans toutes les espèces étudiées jusqu’à présent. Chez les animaux, l’appariement est préférentiellement initié au niveau des télomères. Chez les plantes supérieures et les champignons, chaque bivalent présente plusieurs sites d’initiation. Au pachytène, le complexe est complet dans l’ensemble des bivalents. Sa longueur diminue en même temps que les chromosomes raccourcissent. Le complexe n’est plus visible au diplotène, sauf au niveau des chiasmas où il subsiste dans une forme plus ou moins modifiée jusqu’en diacinèse.Le complexe synaptonémal est donc étroitement lié à l’appariement, et son rôle essentiel est de maintenir à distance égale les deux chromosomes homologues. De plus, dans certains cas de méiose achiasmatique (femelle de ver à soie), il assure ce rôle jusqu’en métaphase. Le complexe est également impliqué dans la recombinaison génétique (cf. infra ): les échanges entre chromatides homologues ont lieu durant sa mise en place dans le noyau, et l’élément central présente une différenciation sous forme de nodule dont la répartition et le nombre correspondent assez bien à ceux du crossing-over. Ces nodules apparaissent au zygotène et disparaissent au diplotène. Leur nombre par noyau correspond au nombre moyen de chiasmas. Alors que femelle et mâle de ver à soie forment des complexes synaptonémaux, seul le mâle présente des crossing-over. Parallèlement, des nodules ne sont observés que chez le mâle.Lorsque le complexe synaptonémal disparaît, le ou les chiasmas sont le seul lien entre les chromosomes homologues. Conséquence de la recombinaison génétique et donc élément caractéristique de la méiose à cause du brassage génétique dont ils sont le reflet, les chiasmas sont aussi un élément indispensable au déroulement correct de la méiose. En effet, si leur nombre varie d’un chromosome à l’autre et d’une méiose à l’autre, leur nombre total par cellule méiotique est toujours supérieur au nombre (n ) des chromosomes de l’espèce. Dans les mutants de méiose où l’on observe une diminution du taux des échanges génétiques, le nombre des chiasmas diminue et certains des chromosomes deviennent achiasmatiques: cela entraîne une mauvaise répartition de ces chromosomes aux stades suivants. Le chiasma a donc un rôle mécanique puisqu’il permet de maintenir les chromosomes homologues jusqu’à ce que les microtubules du fuseau de division (voir fuseau et mitose) prennent le relais. Au cours de la mitose, ce problème ne se pose pas puisque cette division sépare les deux chromatides de chaque chromosome, liées l’une à l’autre au niveau de leur centromère. Durant la métaphase de première division de méiose, ce sont des chromosomes entiers qui sont séparés vers chacun des pôles cellulaires, et l’absence de «liens» entre les homologues conduit à l’indépendance des chromosomes et donc à leur répartition inégale à chaque pôle.Trois événements caractérisent la méiose: l’appariement des chromosomes homologues, leur recombinaison et leur ségrégation. Alors que le lien entre les deux derniers événements est clair (cf. supra ), les résultats concernant le déroulement des deux premiers restent contradictoires. Une analyse détaillée des phases de la méiose chez la levure de boulangerie a permis de préciser les étapes suivantes: la recombinaison est initiée par des cassures doubles-brins de l’ADN distribuées tout au long des chromosomes et souvent localisées dans des gènes connus pour leurs forts taux de recombinaison. Ces cassures se produisent en même temps que le complexe synaptonémal se met en place. Une des hypothèses est de dire que pour réparer ces cassures le chromosome homologue va servir de modèle, ce qui facilitera la recherche des homologues dans le noyau et par conséquent leur appariement. Un seul événement pourrait ainsi être à l’origine de la recombinaison et de l’appariement. La maturation des événements de recombinaison (réparation des coupures et échange des brins) a lieu durant tout le pachytène, vraisemblablement facilitée par la présence du complexe synaptonémal.La métaphase et l’anaphase de première divisionLa métaphase débute avec la formation du fuseau. Les chromosomes sont liés aux fibres fusorales au niveau de leur centromère et se disposent à égale distance des pôles du fuseau sur un plan appelé plan équatorial ou plan métaphasique (fig. 6). Les deux centromères de chaque bivalent se disposent de part et d’autre de ce plan. Cette disposition se fait au hasard: ainsi, chez l’homme dont les méioses comportent 23 paires de chromosomes, il existe 223 associations possibles entre les centromères des chromosomes d’origine paternelle et maternelle entraînant un brassage interchromosomique important.L’anaphase correspond au stade où les deux centromères homologues se séparent, chacun d’eux entraînant deux chromatides à l’un des pôles du fuseau (fig. 6).Les stades télophase et interphase proprement dits n’interviennent pas dans toutes les espèces entre les deux divisions méiotiques. Ces stades impliquent, dans les cas où ils se manifestent, la constitution d’une membrane nucléaire autour des deux noyaux nouvellement formés ainsi que la dédifférenciation des chromosomes de chaque noyau. S’ils sont absents, l’anaphase de première division est immédiatement suivie de la seconde division de méiose. Cependant, dans tous les cas, la première anaphase méiotique a la même conséquence fondamentale: la séparation des centromères homologues entraîne la constitution de deux noyaux qui ne contiennent plus que n centromères mais encore 2 n chromatides (fig. 6).La seconde divisionLa seconde division de la méiose subie par les deux nouveaux «noyaux» est en fait une mitose banale, cependant non précédée d’une duplication de l’ADN. À la métaphase, chacun des n centromères de chaque «noyau» se clive, et les deux chromatides se séparent, permettant la constitution de quatre noyaux à n chromosomes (noyaux haploïdes, fig. 6).Cette description cytologique est très générale. Selon les espèces, on pourrait noter des variantes plus ou moins nombreuses à certains stades, mais dans tous les cas il ne s’agit que de différences de détail ne modifiant en rien les conséquences fondamentales, c’est-à-dire génétiques, de la méiose.Conséquences génétiquesIl serait trop long de résumer ici les nombreux faits d’ordre cytologique, biochimique et génétique qui ont permis d’établir la théorie chromosomique de l’hérédité. On admettra donc simplement que tous les caractères qui définissent un individu sont déterminés par les gènes , unités d’information génétique portés par les chromosomes, et occupant sur ce dernier une place précise ou locus . Si l’information génétique d’un gène est modifiée – on parle de mutation –, le caractère qu’il détermine est lui-même modifié, et cette modification est héréditaire. L’individu mutant diffère donc de l’individu de référence ou individu sauvage . L’un porte un gène muté et l’autre un gène sauvage: ces deux gènes homologues constituent un couple d’allèles .Afin de concrétiser ces notions et d’aborder les lois fondamentales de la génétique, conséquence de la méiose, on considérera les résultats expérimentaux obtenus lorsqu’on réalise un croisement entre individus différant par un ou plusieurs caractères, d’une part chez un organisme à phase haploïde observable, la levure de boulanger (Saccharomyces cerevisiae ), d’autre part chez un organisme où seule la phase diploïde du cycle est observable, la mouche du vinaigre (Drosophila melanogaster ).Ségrégation d’un couple d’allèlesPour qu’un croisement entre deux souches de levure soit possible, c’est-à-dire pour qu’il y ait fécondation, il est nécessaire que les deux souches diffèrent par un couple d’allèles (appelé a / 見). Si cette condition est remplie, on peut réaliser des croisements entre souches présentant de nombreuses différences génétiques. Considérons le cas d’un croisement n’impliquant qu’une seule différence, par exemple entre la souche sauvage, capable de synthétiser la leucine (acide aminé indispensable à sa croissance) et une souche mutante qui, ayant perdu cette propriété par mutation, exige pour sa croissance la présence de leucine dans le milieu de culture. Utilisant un symbolisme classique, on dira que la souche sauvage porte le gène leu + et la souche mutante, son allèle leu . Les résultats d’un tel croisement sont indiqués à la figure 7 ainsi que leur interprétation en termes chromosomiques. La méiose du diploïde leu +/leu produit deux cellules portant le gène leu et incapables de pousser sur milieu sans leucine, et deux cellules portant le gène leu + et capables de pousser sur ce milieu. L’histoire d’un couple d’allèles correspond ainsi à l’histoire d’une paire de chromosomes homologues à la méiose. La séparation des centromères homologues à l’anaphase de première division entraîne une ségrégation 2 : 2 de la paire de chromosomes homologues et par conséquent du couple d’allèles leu / leu +. Pour tout couple d’allèles, une ségrégation 2 : 2 intervient au niveau des produits de la méiose.Chez la drosophile, la souche sauvage présente un corps brun, et l’on connaît, entre autres, une souche mutante dont le corps est noir. Les produits de la méiose sont les gamètes, ovules ou spermatozoïdes selon le sexe, non observables directement. Cependant, le principe de ségrégation 2 : 2 d’un couple d’allèles se manifeste à leur niveau et se répercute au niveau des descendants diploïdes de la génération suivante (tabl. 1). L’observation d’individus diploïdes introduit simplement ici quelques notions génétiques supplémentaires. Chaque mouche diploïde porte dans chacune de ses cellules (à l’exception des gamètes) l’information génétique en double (2 n chromosomes). La présence des gènes b ou b + en deux exemplaires ou celle du gène b sur un chromosome et de son allèle b + sur le chromosome homologue conduit à parler respectivement de génotypes b /b , b +/b + et b +/b . Les individus des deux premiers types sont dits homozygotes , car ils ne donneront naissance qu’à un type génétique de gamète. Un individu de génotype b +/b est hétérozygote : la méiose conduira à deux types génétiques b et b + en proportions égales selon le principe de ségrégation. On peut aussi se demander quel caractère auront les individus hétérozygotes b +/b . Présenteront-ils un corps sauvage (brun) ou un corps noir? Trois cas peuvent se présenter. Si l’hétérozygote (F1) présente le caractère sauvage (on dit qu’il a le phénotype sauvage), cela signifie que le gène b + est dominant sur son allèle b (et que celui-ci est récessif par rapport à son allèle sauvage). Si l’hétérozygote est le phénotype mutant (corps noir), cela signifie que c’est l’allèle mutant qui est dominant sur l’allèle sauvage. Dans les cas où il y a dominance partielle des deux allèles, le phénotype de l’hétérozygote est intermédiaire entre les phénotypes sauvage et mutant [cf. HÉRÉDITÉ].Chez la drosophile, on retrouve le principe de ségrégation 2 : 2 d’un couple d’allèles à la méiose; comme le montre le backcross b +/b 憐 b /b du tableau 1, l’hétérozygote b +/b fabrique deux types de gamètes en proportions égales. Pour le vérifier au niveau de la 2, il suffit de réaliser des backcross individuels entre des individus 2 de phénotype sauvage et des individus homozygotes b /b . On trouve qu’un tiers des individus 2 de phénotype sauvage sont homozygotes b +/b + et deux tiers hétérozygotes b +/b .Le principe de ségrégation illustré par ces deux exemples est l’une des lois génétiques mises en évidence par Mendel dès 1866 lors de son étude sur les pois.Ségrégation indépendante des gènesLa figure 8 illustre et interprète les résultats d’un croisement entre deux souches de levure différant par deux couples d’allèles portés par deux paires de chromosomes. Le comportement des chromosomes à la méiose rend parfaitement compte des résultats observés puisque, lors de la métaphase et de l’anaphase de première division, deux paires de chromosomes ont des histoires indépendantes. En effet, selon leur disposition sur la plaque équatoriale, on obtiendra deux situations à l’anaphase: soit que les deux chromosomes sont de même origine parentale partent ensemble à l’un des pôles; soit que les deux chromosomes d’origine parentale différente partent ensemble vers chaque pôle. Dans le premier cas, à l’issue de la méiose, deux produits présenteront le génotype d’un des parents, et deux, le génotype de l’autre (ce type d’asque ou tétrade est appelé ditype parental ). Dans le second cas, on obtient un asque dont les quatre spores ont un génotype différent de celui des parents: deux sont sauvages et deux doubles mutants. Ces deux génotypes sont dits recombinés, et l’asque est un ditype recombiné . Statistiquement, on constate une égalité quantitative entre les quatre génotypes. Ce type de résultat manifeste la ségrégation indépendante des deux couples d’allèles, et il est la conséquence de l’histoire indépendante des chromosomes à la méiose.Le tableau 2 illustre, chez la drosophile, les conséquences de la production de quatre types de gamètes en proportions égales par un individu 1 double hétérozygote, selon que l’on réalise un croisement par un homozygote récessif (test cross ) ou par un autre hétérozygote 1. Ici aussi, les résultats obtenus en deuxième génération sont la conséquence de l’histoire des chromosomes lors de la méiose chez l’hétérozygote 1.Cette deuxième loi génétique concernant la ségrégation indépendante des gènes fut aussi mise en évidence par Mendel.Liaisons géniquesLorsqu’on réalise un croisement impliquant deux couples d’allèles occupant des loci voisins sur un même chromosome, on n’observe pas une ségrégation indépendante de ces gènes: les génotypes parentaux sont le plus souvent conservés à la méiose. Le tableau 3 donne un exemple de liaison absolue entre deux gènes chez la drosophile. À l’issue de backcross entre des individus mâles de la 1 et des individus femelles doubles mutants, on n’observe que deux catégories d’individus: sauvage et double mutant. Les mâles n’ont donc produit que deux types génétiques de gamètes correspondant aux génotypes parentaux. Dans le cas où ce sont des femelles 1 qui sont croisées par des mâles doubles mutants, on observe un grand excès des deux catégories parentales. L’apparition des types recombinés s’explique par une recombinaison entre les deux loci , à la méiose, au moment où les quatre chromatides sont appariées (prophase de la première division). Ces recombinaisons sont, chez la drosophile, possibles chez la femelle mais inexistantes chez le mâle (cela n’est pas un cas général). Les conséquences de la recombinaison intrachromosomique ou crossing-over sont particulièrement intéressantes à observer chez les organismes où l’on peut analyser les quatre produits de la méiose (cas de la levure). La figure 9 donne un exemple de résultats obtenus lors d’un croisement entre deux gènes liés chez cet organisme. Les différents types d’asques y sont interprétés en termes de crossing-over.On n’a pas envisagé ici la conséquence d’un crossing-over entre le locus d’un gène et son centromère. Cet événement entraîne la formation d’asques tétratypes lors d’un croisement impliquant deux gènes indépendants. La distance gène-centromère peut ainsi être calculée.La fréquence de recombinaison obtenue entre deux loci est caractéristique des couples d’allèles envisagés: elle est reproductible d’un croisement à un autre. Si l’on fait l’hypothèse que plus la distance physique sur le chromosome est grande entre les deux loci , plus la fréquence de recombinaison est élevée, on peut établir une carte génétique des différents gènes connus sur un chromosome. Dans l’exemple indiqué pour la drosophile, on dira que la distance génétique est de 10,7 unités de recombinaison (1 unité = 1 p. 100 de recombinaison), distance calculée en faisant le rapport du nombre de recombinaisons (152 + 154) sur le total d’individus observés (2 840). Tous les gènes liés entre eux constituent un groupe de liaison et, pour une espèce donnée, l’expérience montre que le nombre de groupes de liaisons défini génétiquement correspond toujours au nombre précis de chromosomes observables cytologiquement.Le mécanisme moléculaire de la recombinaison intrachromosomique est encore très mal connu. Il est cependant clair que c’est un processus extrêmement précis qui se produit entre deux chromatides non sœurs d’une paire de chromosomes appariés à la prophase de la première division de la méiose.La méiose apparaît comme un mécanisme adopté par tous les organismes à reproduction sexuée et dont les conséquences les plus fondamentales sont:– La préservation du nombre de chromosomes de l’espèce d’une génération sexuée à la suivante.– La constitution d’une barrière génétique puissante entre les espèces: l’une des causes de la stérilité des hybrides 1 d’espèces est en effet l’absence de synapsis au moment de la méiose. Les deux lots chromosomiques n’ont pas d’homologie suffisante pour qu’il y ait appariement: les gamètes portent des lots chromosomiques déséquilibrés et ne sont pas viables. C’est certainement le cas, par exemple, de la stérilité du mulet, hybride entre l’âne et le cheval.– La production de nouveaux types génétiques. Par le jeu de la ségrégation indépendante des centromères à la première division de la méiose et par les phénomènes de recombinaison intrachromosomiques, la méiose permet la production de génotypes différents de ceux qui existent dans la nature sous le seul effet des mutations et joue ainsi un grand rôle dans l’évolution des espèces.
méiose [ mejoz ] n. f.• 1913; « décours d'une maladie » 1842; gr. meiosis « décroissance »♦ Biol. Division de la cellule en deux étapes aboutissant à la réduction de moitié du nombre des chromosomes contenus dans son noyau (passage du stade diploïde au stade haploïde au cours de la formation des gamètes). ⇒aussi mitose. — Adj. MÉIOTIQUE , 1913 . Réduction méiotique.
● méiose nom féminin (grec meiôsis, diminution) Double division de la cellule aboutissant à la réduction de moitié du nombre des chromosomes, et qui se produit au moment de la formation des cellules reproductrices, ou gamètes. (À l'issue de la méiose, chaque cellule diploïde forme ainsi quatre gamètes haploïdes.)méiosen. f. BIOL Mode de division cellulaire conduisant à une réduction de moitié du nombre de chromosomes de chaque cellule fille.Encycl. La méiose n'affecte que les cellules dont le noyau est diploïde. Elle ne se produit que chez les espèces vivantes soumises à la fécondation, qu'elle précède. Elle comporte deux divisions successives: la première donne deux cellules filles dont le nombre de chromosomes est égal à la moitié de celui de la cellule mère; la seconde est une mitose subie par chacune des deux cellules filles, ce qui donne quatre cellules génétiquement identiques 2 à 2. Lors de la première division, une séparation des gènes allèles s'effectue. La méiose conduit à la formation des gamètes, contenus dans les organes génitaux. La fécondation, en réunissant des gamètes qui ont seulement n chromosomes, donnera à nouveau une cellule à 2n chromosomes. V. mitose.⇒MÉIOSE, subst. fém.BIOL. Dans la formation des gamètes, ensemble des deux divisions cellulaires successives qui terminent l'ovogenèse et la spermatogénèse, et constituant pour la première en une réduction du nombre des chromosomes (synon. division méiotique); en partic. division (réductionnelle) de l'ovogenèse (synon. réduction chromatique, réduction méiotique; division réductrice; mitose réductionnelle). Méiose équationnelle; méiose réductionnelle. [Chez certaines variétés fruitières] la distribution chromosomique à la méiose est irrégulière; pollen et ovules sont anormaux et la germination du pollen est irrégulière (BOULAY, Arboric. et prod. fruit., 1961, p.45). Dès 1906, Yamanouchi décelait la méiose chez une algue rouge (Hist. gén. sc., t.3, vol.2, 1964, p.792). Au moment de la méiose, par suite du phénomène de la réduction, chacun des gamètes reçoit l'un des deux éléments de la paire chromosomique et n'en reçoit qu'un seul (M. LAMY, Les Maladies héréditaires, Paris, P.U.F., 1970, n°1177, p.45). V. mitose ex. de Husson, Graf.Prononc.:[mejo:z]. Étymol. et Hist. 1840 (Ac. Compl. 1842: Meiose (didact.) Déclin, décroissement, diminution); 1913 biol. (CAULLERY, Les problèmes de la sexualité, Paris, Flammarion, p.24-26 ds QUEM. DDL t.22). Empr. au gr.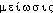 «amoindrissement, diminution». Le mot gr. est également passé en angl. où il est att. comme terme de rhét. dès la fin du XVIe s. et proposé d'abord sous la forme maiosis en 1905 (FARMER et MOORE ds NED Suppl.2), comme terme de biologie.DÉR. Méiotique, adj. Qui concerne la méiose. Division, réduction méiotique (des chromosomes). Lillie (...) a cherché à analyser les forces en présence au cours des divisions mitotiques et méiotiques: attractions et répulsions sont alors utilisées, à bon escient, les centromères étant pris à contribution au même titre que les pôles (Biol., 1965, p.245 [Encyclop. de la Pléiade]). — [
«amoindrissement, diminution». Le mot gr. est également passé en angl. où il est att. comme terme de rhét. dès la fin du XVIe s. et proposé d'abord sous la forme maiosis en 1905 (FARMER et MOORE ds NED Suppl.2), comme terme de biologie.DÉR. Méiotique, adj. Qui concerne la méiose. Division, réduction méiotique (des chromosomes). Lillie (...) a cherché à analyser les forces en présence au cours des divisions mitotiques et méiotiques: attractions et répulsions sont alors utilisées, à bon escient, les centromères étant pris à contribution au même titre que les pôles (Biol., 1965, p.245 [Encyclop. de la Pléiade]). — [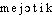 ]. — 1re attest. 1913 (CAULLERY, loc. cit.); de méiose, suff. -ique selon le dér. gr.
]. — 1re attest. 1913 (CAULLERY, loc. cit.); de méiose, suff. -ique selon le dér. gr. 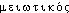 et prob. d'apr. l'angl. meiotic attesté comme terme de biol. dep. 1905 (NED Suppl.2).méiose [mejoz] n. f.ÉTYM. 1913; cf. v. 1910, angl. meiose; en 1842, « décours d'une maladie »; grec meiosis « décroissance ».❖♦ Biol. Division de la cellule (⇒ Mitose) en deux étapes aboutissant à la réduction de moitié du nombre des chromosomes contenus dans son noyau (passage du stade diploïde au stade haploïde au cours de la formation des gamètes).REM. Le mot désigne habituellement l'ensemble des deux divisions que comporte la formation des gamètes (ovogenèse et spermatogenèse), la première étant hétérotypique et consistant seule en une réduction (la seconde est homéotypique et équationnelle).1 Lorsque l'organisme adulte et sexuellement mûr produit les cellules génératrices, c'est un nouveau mécanisme qui intervient, et non moins précis que la mitose, mais dont l'effet est de réduire de moitié le nombre des chromosomes. Au lieu de recevoir le double ensemble de chromosomes qui appartient à toutes les cellules du corps (ou cellules somatiques), chaque cellule génératrice n'en reçoit qu'un seul ensemble où figure un seul chromosome de chaque sorte, lequel peut être ou le paternel ou le maternel (…) si cette réduction chromatique (ou méiose) n'avait pas lieu, le nombre total des chromosomes irait en doublant à chaque génération : la réduction chromatique, en composant le doublement chromosomique résultant de la fécondation, assure la constance numérique des chromosomes (…)Jean Rostand, Idées nouvelles de la génétique, p. 9.2 Réduction chromatique ou méiose. L'oogenèse et la spermatogenèse offrent, dans les deux divisions cellulaires qui les terminent, un processus général et particulièrement important, mis en évidence à la fin du XIXe et au début du XXe siècle, la réduction chromatique, ou méiose (…)Maurice Caullery, l'Embryologie, p. 14-15.♦ Spécialt. La réduction chromosomique proprement dite (première division).❖DÉR. Méiotique.
et prob. d'apr. l'angl. meiotic attesté comme terme de biol. dep. 1905 (NED Suppl.2).méiose [mejoz] n. f.ÉTYM. 1913; cf. v. 1910, angl. meiose; en 1842, « décours d'une maladie »; grec meiosis « décroissance ».❖♦ Biol. Division de la cellule (⇒ Mitose) en deux étapes aboutissant à la réduction de moitié du nombre des chromosomes contenus dans son noyau (passage du stade diploïde au stade haploïde au cours de la formation des gamètes).REM. Le mot désigne habituellement l'ensemble des deux divisions que comporte la formation des gamètes (ovogenèse et spermatogenèse), la première étant hétérotypique et consistant seule en une réduction (la seconde est homéotypique et équationnelle).1 Lorsque l'organisme adulte et sexuellement mûr produit les cellules génératrices, c'est un nouveau mécanisme qui intervient, et non moins précis que la mitose, mais dont l'effet est de réduire de moitié le nombre des chromosomes. Au lieu de recevoir le double ensemble de chromosomes qui appartient à toutes les cellules du corps (ou cellules somatiques), chaque cellule génératrice n'en reçoit qu'un seul ensemble où figure un seul chromosome de chaque sorte, lequel peut être ou le paternel ou le maternel (…) si cette réduction chromatique (ou méiose) n'avait pas lieu, le nombre total des chromosomes irait en doublant à chaque génération : la réduction chromatique, en composant le doublement chromosomique résultant de la fécondation, assure la constance numérique des chromosomes (…)Jean Rostand, Idées nouvelles de la génétique, p. 9.2 Réduction chromatique ou méiose. L'oogenèse et la spermatogenèse offrent, dans les deux divisions cellulaires qui les terminent, un processus général et particulièrement important, mis en évidence à la fin du XIXe et au début du XXe siècle, la réduction chromatique, ou méiose (…)Maurice Caullery, l'Embryologie, p. 14-15.♦ Spécialt. La réduction chromosomique proprement dite (première division).❖DÉR. Méiotique.
Encyclopédie Universelle. 2012.